Extracellular Matrix and musculoskeletal systems
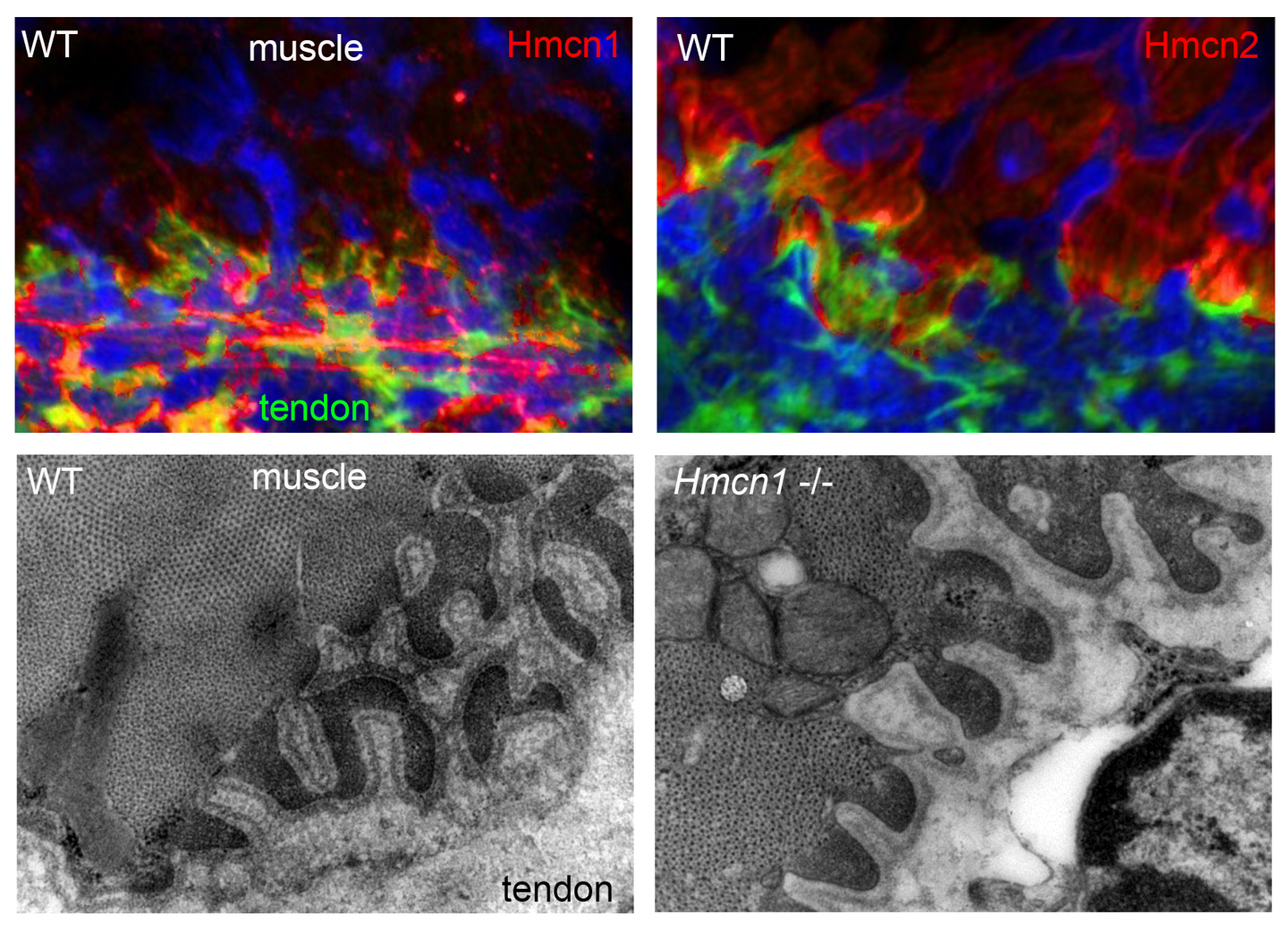
Based on our forward genetic screens in zebrafish, we had identified several extracellular matrix proteins, such as the Fibulin family members Hmcn1, Hmcn2 and Fibulin-1, as well as Fibrillin-2, a known component of microfibers, as essential for proper epidermal-dermal junction formation (leading to embryonic skin blistering upon mutation; Carney et al., PLoS Genet. 2010). To elucidate the role of Hmcn1 and its close relative Hmcn2 in mammals, we have raised specific antibodies for comparative protein distribution studies and have knocked them out in the mouse. However, mutants do not display blistering defects and are viable and fertile. At the ultrastructural level (revealed via transmission electron microscopy), adult Hmcn1 mouse mutants display compromised epidermal-dermal and myotendinous junctions, which, however, do not affect overall skin and skeletal muscle anatomy and physiology (Welcker et al., Sci. Rep. 2021). Loss of Hmcn2, on the other hand, seems to lead to compromised endurance of adult mice in grip strength tests. A similar excitation-induced muscle fatique has been reported as a symptom of tubular aggregate myopathies caused by mutations in genes involved in T-tubule-based excitation-contraction coupling. Of note, such T-tubules of skeletal muscle, which according to our analyses contain high amounts of Hmcn2 protein, are intracellular invaginations of the cell membranes of skeletal myotubes together with their attached basement membranes, thus constituting a structure with two juxtaposed basement membranes as in myotendinous junctions that are affected in Hmcn1 mutant mice (Welcker et al., Sci. Rep. 2021) and as in epidermal-dermal-epidermal and epidermal-dermal-myomuscular junctions affected in zebrafish Hmcn1 and Hmcn2 mutations, respectively (Carney et al., PLoS Genet. 2010; Feitosa et al., Dev. Biol. 2016). Together, this suggests that similar pathomechanisms might underlie the muscular fatique defects observed in mouse Hmcn2 mutants, a notion that is currently further investigated. Furthermore, we are looking into potential roles of Hmcn2 in myomuscular junctions in the myosepta between somites of zebrafish embryos.
In addition, as a first step to unravel the molecular mechanisms by which Hmcn proteins contribute to proper linkage of epithelia (and underlying basement membranes) to surrounding tissues, we have applied biochemical approaches (e.g. Surface Plasmon Resonance Spectroscopy) to identify potential physical binding partners of mouse and zebrafish Hmcn1. These studies are carried out in cooperation with Prof. Gerhard Sengle from the Institute for Biochemistry II of the University Hospital Cologne, and are part of the DFG-funded Research Unit FOR2722. One identified binding partner is Fibrillin-1 (unpublished data), in line with our former finding that during embryonic blistering in zebrafish embryos, Hmcn1 displays a genetic interaction with Fibrillin-2 (Carney et al., PLoS Genet. 2010). Another identified novel binding partner is Nidogen-2, an integral component of basement membranes, explaining how Hemicentin- and Fibrillin-based microfibers of the extracellular matrix are linked to epithelial basement membranes both in skin and in skeletal muscle (Zhang et al., Matrix Biol. 2022).